2022年6月2日,福建农林大学海峡联合研究院基因组中心张积森团队在《自然-遗传学》(Nature Genetics)上发表了题为“Genomic insights into the recent chromosome reduction of autopolyploid sugarcane Saccharum spontaneum”的研究论文,这是基因组中心团队继2018年解析甘蔗基因组(Zhang et al., 2018, Nature Genetics)后的又一重大进展。该研究解析了细茎野生种天然同源四倍体Np-X基因组,并利用基因组学手段系统阐明了甘蔗细茎野生种(又称割手密)的起源、染色体基数、基因组倍体、关键性状相关基因的演化,为甘蔗的基因组辅助育种奠定了重要的理论基础。

现代栽培甘蔗是100多年前高贵种与细茎野生种人工杂交后产生,高贵种是同源八倍体,种内的材料拥有稳定的核型(2n=8x=80),而细茎野生种为现代甘蔗栽培种贡献了抗逆、分蘖等优良性状,也实现了甘蔗在育种史上最大的突破。但细茎野生种分布区域广(从地中海到太平洋),并且其具有宽泛的倍性变异(从四倍到十六倍)和基础染色体数变异(8,9,10),其中,十六倍体的细茎野生种是甘蔗属已知的倍性最高的材料,由于细茎野生种遗传背景的高度复杂,其起源和演化是研究界百余年来悬而未决的重要科学问题,这极大地限制了现代甘蔗育种的进程。
甘蔗属的遗传背景极其复杂,最早有关甘蔗属染色体核型的报道可以追溯到1929年,Bremer用经典细胞学鉴定了杂种甘蔗POJ2725的染色体数为106-107,但是鉴定结果极不稳定。随着科技的进步,以经典细胞学为基础衍生出来的分子细胞学为甘蔗遗传背景的鉴定提供了重要的手段,但由于实验的手法限制,并且甘蔗是复杂的多倍体,诸多的干扰因素仍然给甘蔗属遗传背景的解析带来了大量困难,因此,百余年来,甘蔗属的遗传背景一直是未解之谜。自人类基因组计划完成以来,基因组测序技术得到了飞速发展,为复杂物种遗传背景的解析带来了新的契机。研究团队突破了同源多倍体基因组组装的挑战,成功解析了甘蔗细茎野生种遗传背景的演化规律,将为甘蔗的遗传育种提供重要的理论指导。
一、一个祖先核型同源多倍体细茎野生种基因组组装与注释。
研究选择了染色体基数为x = 10的自然同源四倍体割手密Np-X(2n = 4x = 40)为对象进行了深入研究。为了克服同源多倍体分型组装上的困难,研究人员利用高准确性的基于PacBio测序平台的CCS reads对其进行了基因组组装,并利用Hi-C技术对该基因组进行了挂载,最终得到了2.76 Gb染色体水平的基因组,并且在超过92%的染色体中组装出了着丝粒结构。为了确保同源组内分型的准确,研究还利用了基于ONT平台的Ultra-long reads对基因组的组装质量进行了独立验证,超过90%的Ultra-long reads能够以高匹配度比对到组装的基因组上,并且同源染色体之间的错误连接率不足4%。高质量的Np-X基因组的获得为基因组结构和群体基因组学分析奠定了重要的基础。
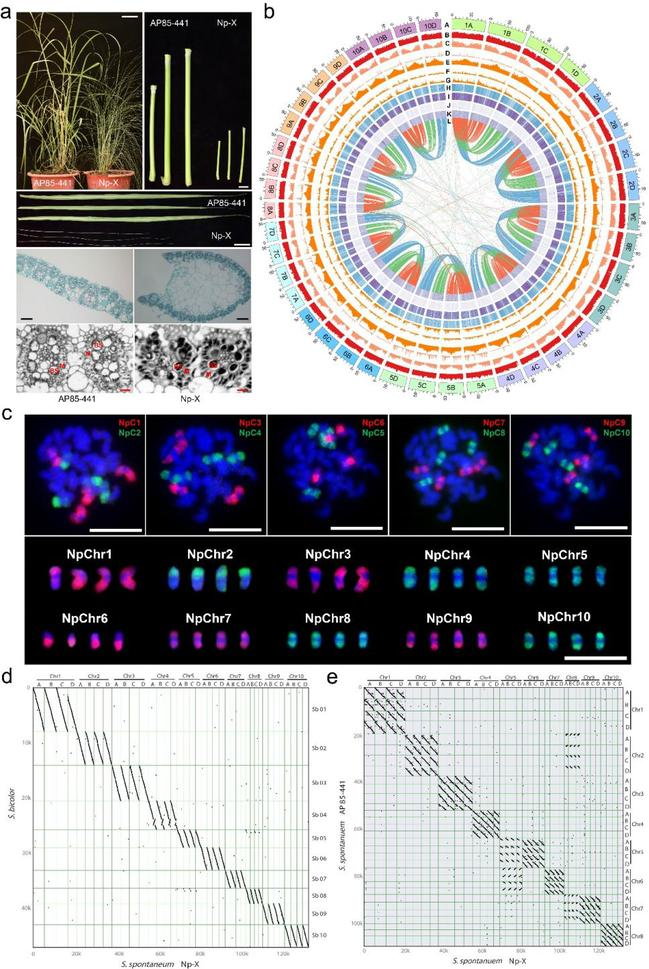
图1. 细茎野生种Np-X与AP85-441的表型、核型和基因组特征的比较
a. 细茎野生种Np-X与AP85-441表型的比较;b. Np-X的基因组特征;c. Np-X的核型;d. Np-X与高粱的共线性比较;e. Np-X与AP-441的共线性比较。
二、一对古复制的染色体的近期演化。
通过基因组结构的比较,结合FISH证据和HiC信号的结果发现了割手密种染色体基数由x = 10到x = 8的演化是由于祖先的5号和8号染色体在着丝粒区域发生了断裂,断裂的着丝粒在重组后的染色体上逐渐退化,并且,研究人员在断裂的着丝粒上分别鉴定到了6.5Mb和7.1Mb区域的丢失,并在该区域分别鉴定到了15和28个基因。
祖先的5号和8号染色体是来源于禾本科ρ事件的一对古复制的染色体,这两条染色体上存在着大量的抗病相关的R基因(Zhang et al., 2018),意味着这两条染色体极具育种利用的潜力,为了理清这两条染色体的演化特征,研究人员从整个禾本科的视角出发,探究它们的协同演化规律,发现这两条染色体上的基因在x = 8的AP85-441中相对于x = 10的Np-X受到了更弱的选择。通过转录组发现这两条染色体上的基因存在大量的剂量冗余,说明它们正在进行快速的进化。
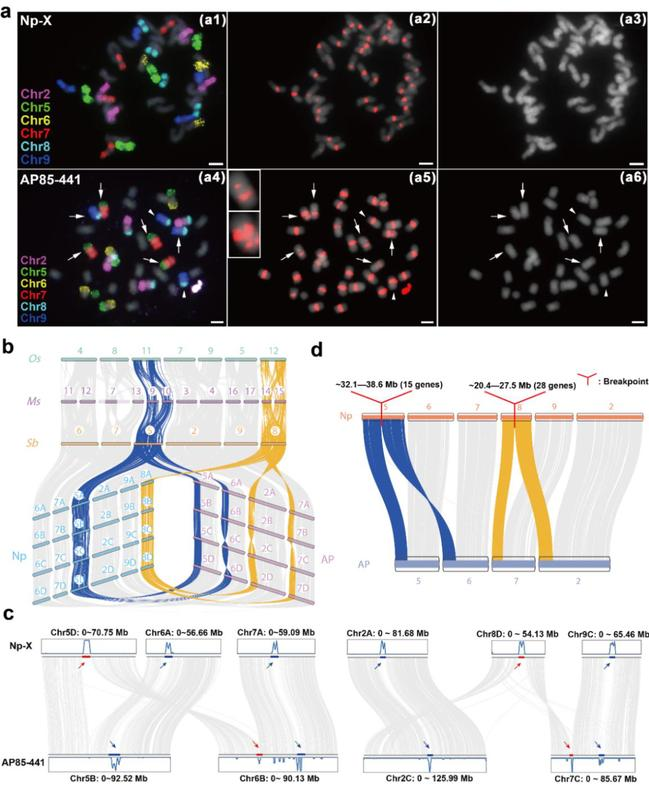
图2. 古复制的染色体对的演化
a. Oligo-Fish分析割手密Np-X与AP85-441的中期染色体;b. 古复制的染色体对在禾本科的演化;c. 古复制的染色体对在AP85-441中发生了染色体重排;d. 重组断点区域丢失的基因。
三、甘蔗属关键生物学分化性状的基因演化。
研究鉴定了多个甘蔗关键性状(糖分积累、糖分代谢、叶片宽窄、光合作用、抗病等)相关的家族基因,并解析了其在禾本科中的系统演化关系。研究基于甘蔗属内的表达谱找到8个基因可能与细茎野生种Np-X的光合作用相关,并发现甘蔗属内参与C4途径的成员基因的类型和其受到的表达调控表现出趋同进化。研究还发现糖转运蛋白基因PLT家族在Np-X和AP85-441基因组中均发生了扩张,而STP基因家族的扩张只发生AP85-441基因组中,这些基因家族的成员可能在这两个细茎野生种材料中是独立演化的。研究比较了热带种和细茎野生种甘蔗糖分转运蛋白基因的上游调控序列的结构,认为在甘蔗属中调控糖分积累的机制是依靠调控转录表达进行的,而不是通过功能基因的获得或丢失。此外,研究还利用热带种、细茎野生种的转录组与叶片宽度的性状进行关联,鉴定到了2个与叶片宽度相关的NAL基因(NAL1和NAL10),可以用于后续的功能验证。
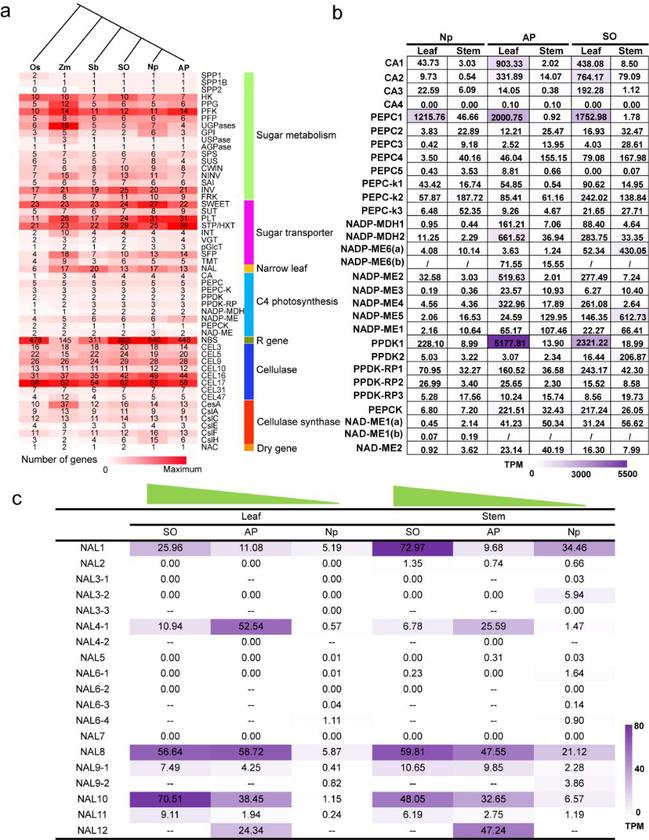
图3. 甘蔗重要性状相关基因家族的扩张
a. 甘蔗重要性状相关基因家族成员在禾本科中的分布;b. C4光合相关基因在三个甘蔗材料不同组织中的表达谱;c. 控制叶片宽窄性状的关键基因在三个甘蔗材料不同组织中的表达谱。
四、甘蔗属的多倍化与演化。
为了理清甘蔗属的系统演化关系,研究人员分别利用碱基同义替换率和LTR类型的转座子插入事件对甘蔗属内的演化规律进行了解析,发现割手密种与热带种的分化时间为1.6百万年,割手密Np-X与AP85-441的分化时间为0.8百万年。明确了以x = 10的割手密核型是甘蔗属核型的祖先模式,并且八倍体的热带种(2n = 8x = 80)和割手密种(2n = 8x = 64)都经历了两次全基因组加倍事件,而四倍体的割手密Np-X只经历了一次加倍事件。
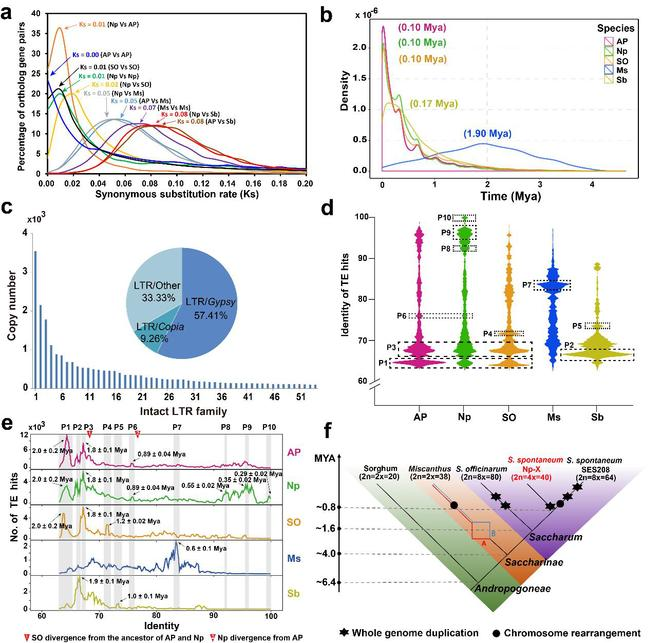
图4. 甘蔗属的演化
a. 甘蔗属及其近缘种间的碱基同义替换率(Ks);b. 甘蔗属及其近缘种转座子的爆发时间;c. Np-X基因组中全长LTR-RTs的分类;d. 甘蔗属中转座子序列一致性分析;e. 甘蔗属中不同序列一致性转座子峰值的时间估计;f. 甘蔗属演化结构。
五、细茎野生种种群的染色体基数与倍性的近期演化。
甘蔗细茎野生种具有非常宽泛的遗传背景和广泛的地理分布,细茎野生种被用于甘蔗育种已有百余年的历史,然而,细茎野生种的起源和群体演化一直是个谜,为了研究细茎野生种群体的遗传多样性,研究人员基于高质量的细茎野生种Np-X基因组,对分布在世界范围内的102份割手密材料进行了遗传多样性分析,系统进化树和群体结构分析表明细茎野生种起源于印度北部,细茎野生种群体可以分为4个亚群,其中,亚群1与亚群2之间存在较弱的基因交流,这4个亚群之间独立演化。
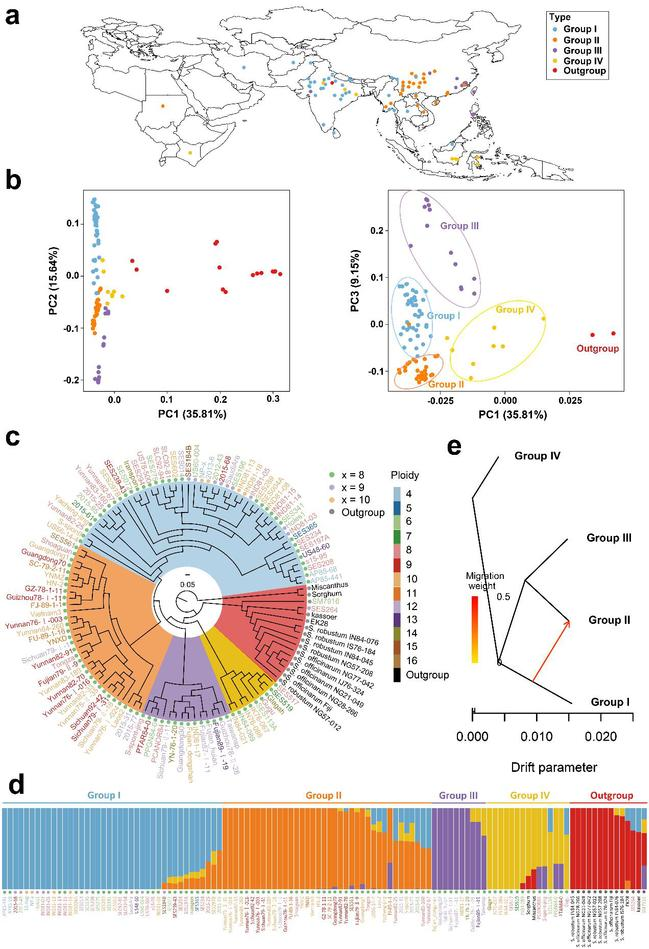
图5. 细茎野生种群体的遗传关系
a. 细茎野生种重测序样本地理位置分布; b. PCA分析; c. 系统发育树分析; d. 群体结构分析; e. 基因流分析。
染色体基数变异广泛存在于甘蔗细茎野生种中,传统依赖于细胞学FISH手段鉴定细茎野生种染色体基数的方法固然有效,但是存在实验误差和探针非特异性结合等因素的干扰,并且实验周期较长。为此,研究人员开发了一种基于5号和8号染色体着丝粒区域reads覆盖度的方法来鉴定细茎野生种的染色体基数,该方法快捷、简易、准确、高效,为甘蔗遗传背景的鉴定提供了重要的策略。此外,研究人员通过对染色体基数为x = 8,x = 9 和x = 10的细茎野生种群体进行遗传多样性分析揭示了细茎野生种的倍性是独立演化的,并且x = 8的群体没有经历人工选择。通过对细茎野生种有效群体大小分析表明,细茎野生种群体在12 ~ 14万年前的有效群体大小达到了最大值,而在0.8 ~ 1.4万年前经历了群体瓶颈效应,可能与当时全球气候经历“新仙女木”事件有关。最后,该研究系统总结了细茎野生种群体的演化规律,并提供了细茎野生种遗传背景的演化模型。
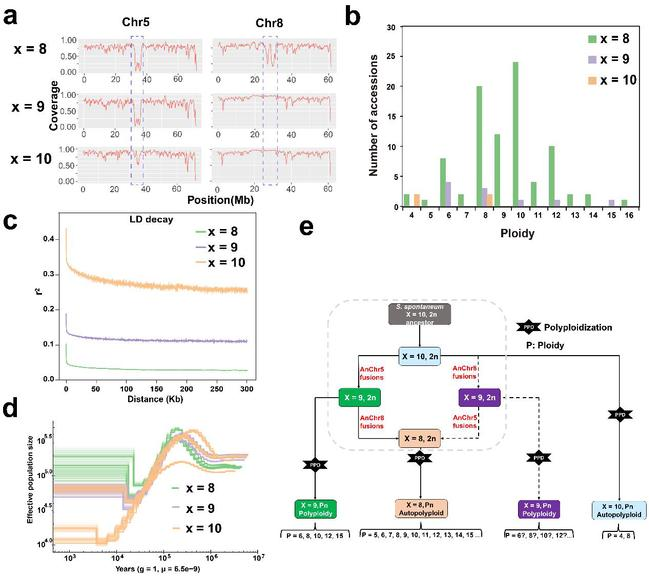
图6. 割手密群体的进化
a. 基于二代reads比对的覆盖度对不同染色体基数进行判定;b. 102份细茎野生种重测序材料的遗传背景;c. 不同染色体基数亚群的连锁不平衡分析;d. 割手密群体的种群演化历史;e.割手密不同染色体基数的演化。
研究通过利用基因组学手段系统阐明了甘蔗细茎野生种的起源、遗传背景及群体历史,深入挖掘了关键性状相关基因的演化,将会全面加强甘蔗野生种种植资源的保护与利用和推动甘蔗全基因组辅助育种。
福建农林大学海峡联合研究院基因组中心张积森课题组博士生张清(目前在中国农科院深圳农业基因组研究所工作)、齐浥颖、硕士生潘浩然和基因组中心唐海宝教授为论文共同第一作者,基因组中心张积森教授为通讯作者。张积森课题组已毕业博士王刚、华秀婷(目前在广西大学工作)、博士生王勇军、李珍、王天友、马盼盼、硕士生李艺寒、窦梅杰、唐海宝课题组硕士生王毅斌、广西大学张木清教授、陈保善教授、姚伟副教授、余凡博士、硕士生于泽怀、福建农林大学刘仲健教授、邓祖湖研究员、徐景升教授、王恒波博士、闽江学院黄永吉博士、中国农业科学院深圳农业基因组所张兴坦研究员、云南农业大学杨清辉教授、华中农业大学王茂军教授、云南甘蔗研究所刘新龙研究员、佛罗里达大学王建平教授和伊利诺伊大学香槟校区明瑞光教授为参与作者。该研究得到了国家重点研发计划、广东省科技计划、国家自然科学基金、原国家863计划、福建农林大学优秀博士论文基金、国家留学基金委、中国博士后基金等项目的联合资助。
张积森教授团队长期从事甘蔗基因组学与分子育种研究,该成果是基因组中心团队在甘蔗领域发表的第二篇《自然遗传学》研究论文,这两篇研究论文是近60年以来甘蔗研究领域发表的刊物级别最高的论文,奠定了我校在甘蔗基础研究领域的国际领先学术地位。
文章链接:
https://www.nature.com/articles/s41588-022-01084-1
附:评论二则
田志喜 研究员(中国科学院遗传与发育生物学研究所)
多倍体一般分为两大类:同源多倍体和异源多倍体。普遍认为,同源多倍体是由同一物种内的全套染色体的复制产生的。由于多倍体的基因组分析极具挑战性,尤其是同源多倍体,因此对同源多倍体基因组进化的研究与异源多倍体相比要少得多。甘蔗细茎野生种染色体数目变化范围可从40条到128条,这种广泛的差异与其倍性水平(4x-16x)和基础染色体数(x = 8, 9, 10)有关,是研究多倍体进化的优秀材料。
福建农林大学海峡联合研究院基因组中心张积森团队长期开展甘蔗基因组学等基础生物学研究。2018年该团队解析了一个同源八倍体甘蔗细茎野生种SES208(2n= 8x = 64)的单倍体基因组——AP85-441(1n= 4x = 32),发现其基础染色体数为8,相关结果发表在《自然-遗传学》期刊。虽然该工作开创性解决了在同源多倍体中解析等位基因这一具有挑战性的技术问题,但关于同源多倍体在甘蔗属中是如何进化的关键问题仍不清楚。在本研究中,作者组装了一个基础染色体数为10的野生同源四倍体甘蔗细茎野生种Np-X基因组(2n = 4x = 40),该基因组代表了野生甘蔗的祖先核型。在此基础上,研究团队收集并重测序了116份甘蔗材料(测序总数据量为4682 Gb),探究了细茎野生种的起源和确切的染色体演化模式。该研究发现:
1. 通过对细茎野生种Np-X和细茎野生种AP85-441的比较基因组学分析,特别是与古复制染色体对(PdCPs)相关的基因组学比较,作者发现甘蔗染色体不同倍性的演变是经过了裂变和融合的过程,重组断点位于NpChr5和NpChr8的着丝粒上,该演变过程大约发生在80万年前。
2. 通过基因组3D结构分析,作者有意思地发现A区室(活性区)/ B区室(非活性区)分类在同源染色体上的等位基因之间并不是完全保守的,暗示在同源多倍体中,同源染色体之间虽然高度共线,但染色质结构可能存在高度多样化。此外还发现,在细茎野生种中,PdCPs的位点可能抑制了染色质从非活性状态向活性状态的转换,表明染色体数目的减少可能在基因调控中发挥了重要的作用。
3. 根据群体遗传学的分析结果,作者将收集到的细茎野生种材料分为了4个亚群,这4个亚群的地理位置从印度次大陆到东亚和南亚连续分布。这4个亚群起源于多样性中心印度(group1),随后扩散到了group2,group3,group4亚群所在的地区。group1所含样本的基础染色体数包括x = 8、9、10三种类型;group2,group3,group4亚群中的样本的基础染色体数都以x = 8为主。结合reads比对的方法,作者推断出甘蔗基础染色体数的变化是一个逐步的过程,即从x = 10到x = 9再到x = 8,明确了甘蔗的染色体演化模式。
4. 此外,作者还发现在甘蔗细茎野生种中,三种不同基础染色体数的亚群独立地从祖先进化。大约在120~ 140 Kya,细茎野生种群体经历了有效群体(Ne)的扩张(Ne ~ 500,000),随后在8 ~ 14 Kya又经历了Ne收缩(Ne ~ 10,000~ 60,000)。作者鉴定到一些在各亚群之间表现出差异的选择清除区域,这些区域可能与染色体减少过程中的多糖代谢和胁迫耐受相关。在现代甘蔗品种中,这种适应性可能有助于塑造甘蔗细茎野生种作为逆境抗性遗传供体的角色。
5. 作者对甘蔗重要农艺性状相关的基因家族进行了分析,发现与糖积累相关的基因家族中,INV基因的扩张可能是高糖甘蔗进化的先决条件;并且在Np-X和AP85-441两个细茎野生种中,与C4光合途径高度相关的NAD-ME基因家族成员均发生了扩张,但在高贵种中没有此发现。此外,作者还发现了可能解释不同甘蔗种质间叶片宽度差异的功能候选基因。该分析为解析甘蔗关键农艺性状的遗传机制奠定了重要的基础。
该研究首次系统解析了甘蔗细茎野生种种质资源的演化,极大地提高了对同源多倍体基因组进化以及甘蔗生物学的认识,是甘蔗基础研究领域的一个里程碑式的进展,对甘蔗从头杂交育种具有十分重要的理论指导意义。该研究也为多倍体植物的演化提供了先驱性的研究榜样,引领未来多倍体植物的基因学研究。
黄学辉 教授(上海师范大学)
与小麦、棉花、香蕉等百万年前产生的天然多倍体不同,现代栽培种甘蔗是在一百年前由八倍体的热带种(2n = 8x = 80)与细茎野生种甘蔗反复杂交产生的。热带种甘蔗核型固定并且遗传背景狭窄,因而目前难以从热带种种质资源中去拓展现代栽培种甘蔗的遗传多样性。细茎野生种甘蔗地理分布广泛,为现代栽培种甘蔗贡献了抗逆、分蘖等优良性状。甘蔗细茎野生种倍性和染色体基数的广泛变异,是甘蔗育种最为重要的种质资源,为研究植物同源多倍体基因组的进化提供了一个极为有趣的模型。
福建农林大学张积森课题组组装了甘蔗细茎野生种天然同源多倍体基因组。由于同源多倍体的分型组装至今还是一个基因组研究的难题,因而作者在分型组装的基础上,还基于Ultra-Long reads以分段滑窗方式对基因组组装质量进行了独立验证,这是对现有多倍体基因组组装策略的创新性补充。细茎野生种甘蔗存在三种基础染色体数:x = 8, x = 9,x = 10。通过比较基因组学分析,作者证实了他们组装的Np-X(x = 10)是保持祖先核型的甘蔗细茎野生种。
应用这个祖先核型的基因组,他们揭示了一系列有意思的“近期”演化事件:
1)甘蔗细茎野生种的染色体分裂整合的断点是在着丝粒区域,并且着丝粒再重组过程是带有着丝粒结构的染色体片段融合入另外一条完整的染色体,导致x = 8的细茎野生种出现“双着丝粒”现象。着丝粒序列高度复杂,该项工作可能是首次在植物中揭示了“双着丝粒”的演化事件。
2)重组的染色体是一对ρ事件复制的古染色体对,存在剂量冗余,其中5号染色体上的基因表达相对于其他染色体明显受到了抑制。而该染色体对上存在大量的NBS-LRR基因,并且这对古复制的染色体对在水稻、高粱等作物中也完整保留着,在其他作物中是否存在类似的现象是禾本基因组学的重要科学问题。
3)种内重组的染色体会促进失活染色体区室(B-compartment)的形成,对植物种内“近期”演化的研究具有重要的参考价值。
4)甘蔗属的分化低于1.6百万年,其多倍体也是“近期”事件,约为0.8百万年内。此外,作者对甘蔗属两个核心原始种——细茎野生种和热带种进行了系统的基因组学比较,鉴定了一系列两个甘蔗种“近期”独立分化的且与糖分代谢、生物量、光合等相关的基因,为甘蔗的分子辅助育种提供了重要的理论指导。
细茎野生种在染色体数目上具有很大的差异,从2n = 40到2n = 128不等,基因组倍性水平也从4倍到16倍不等。作者通过100多份代表性的细茎野生种材料的群体基因组学研究,揭示了细茎野生种的倍体是独立起源的,提出印度北部是甘蔗细茎野生种的起源中心;作者巧妙应用祖先核型的染色体比对二代测序数据覆盖特征,鉴定了群体中x = 8, x = 9,x = 10的材料;并阐明了甘蔗细茎野生种是经过5号染色体先断裂,再经8号染色体的演化和整体多倍体化的过程;对基础研究x = 8的群体环境适应性的优势做了基因组学解析。这一系列研究,有望为甘蔗种质资源的开发和利用提供理论指导。
此外,多倍体化是植物进化的重要动力之一,多达70%的开花植物是多倍体化后不久产生的。由于已发布的同源染色体水平的基因组较少,之前基因组信息的缺乏阻碍了同源多倍体基因组进化的研究。张积森课题组的甘蔗细茎野生种研究是同源多倍体研究的先驱工作,将引领多倍体基因组学的发展。
